How to Read Dna Molecular Weght Markers

A molecular-weight size marker in the form of a 1kb Dna ladder in the rightmost lane, used in gel electrophoresis. Gel conditions are 1% agarose, 3 volt/cm, and ethidium bromide stain.
A molecular-weight size marker, as well referred to equally a protein ladder, Dna ladder, or RNA ladder, is a set of standards that are used to place the approximate size of a molecule run on a gel during electrophoresis, using the principle that molecular weight is inversely proportional to migration rate through a gel matrix. Therefore, when used in gel electrophoresis, markers finer provide a logarithmic scale by which to approximate the size of the other fragments (providing the fragment sizes of the mark are known).
Protein, DNA, and RNA markers with pre-determined fragment sizes and concentrations are commercially available. These can be run in either agarose or polyacrylamide gels. The markers are loaded in lanes side by side to sample lanes earlier the first of the run.
DNA markers [edit]

Electrophoresed gel with DNA ladders of varying lengths in left lane and middle lane. Fragments sizes are marked on the right, in base pairs.
Development [edit]
Although the concept of molecular-weight markers has been retained, techniques of evolution have varied throughout the years. New inventions of molecular-weight markers are distributed in kits specific to the marking's type.
An early on problem in the development of markers was achieving high resolution throughout the entire length of the mark.[1] Depending on the running conditions of gel electrophoresis, fragments may accept been compressed, disrupting clarity. To address this issue, a kit for Southern Blot assay was developed in 1990, providing the first marker to combine target DNA and probe Deoxyribonucleic acid. This technique took advantage of logarithmic spacing, and could exist used to identify target bands ranging over a length of 20,000 nucleotides.[2]
Design [edit]
There are 2 common methods in which to construct a Deoxyribonucleic acid molecular-weight size marker.[iii] I such method employs the technique of partial ligation.[three] DNA ligation is the process past which linear DNA pieces are connected to each other via covalent bonds; more specifically, these bonds are phosphodiester bonds.[iv] Hither, a 100bp duplex DNA slice is partially ligated. The outcome of this is that dimers of 200bp, trimers of 300bp, tetramers of 400bp, pentamers of 500bp, etc. will form. Additionally, a portion of the 100bp dsDNA will remain. Equally a effect, a DNA "ladder" equanimous of Deoxyribonucleic acid pieces of known molecular mass is created on the gel.[three]
The second method employs the use of restriction enzymes and a recognized DNA sequence.[3] The DNA is digested by a particular restriction enzyme, resulting in DNA pieces of varying molecular masses. One of the advantages of this method is that more marker tin readily exist created only by digesting more than of the known Deoxyribonucleic acid.[3] On the other manus, the size of the Dna pieces are based on the sites where the restriction enzyme cuts. This makes it more hard to control the size of the fragments in the marking.[5]
More than recently, another method for constructing DNA molecular-weight size markers is beingness employed by laboratories. This strategy involves the use of Polymerase Chain Reaction (PCR).[5] This is achieved one or two means: i) a DNA target is amplified at the same fourth dimension via primer sets, or 2) different Dna targets are amplified independently via particular primers.[5]
Furnishings of gel conditions [edit]
Every bit with experimental samples, the conditions of the gel can bear upon the molecular-weight size mark that runs alongside them. Factors such every bit buffer, charge/voltage, and concentration of gel tin affect the mobility and/or advent of your marker/ladder/standard. These elements need to be taken into consideration when selecting a marker and when analyzing the terminal results on a gel.

1.2% agarose gel showing two different Dna ladders dyed with GelRed stain
- Buffers
- Buffers act to 1) establish pH, and two) provide ions to back up electrical conductivity. In DNA electrophoresis, the TAE (Tris-acetate-EDTA) and TBE (Tris-borate-EDTA) are the usual buffers of pick.[6] TBE buffer is preferred for small Dna pieces, whereas TAE is better suited for fragments greater than 1500 base pairs. In terms of buffering chapters, TAE is lower when compared to TBE; this generally results in slower mobility of the DNA. TBE is also capable of meliorate resolution.[7]
- It must be noted that water cannot act as a substitute for one of these buffers, as the DNA will non drift along the gel.[6] Furthermore, using water instead of buffer will effect in the gel melting.[8]
- Accuse/Voltage
- In terms of voltage, the recommended range is between 4 and 10 5/cm (i.e., volts/cm).[viii] Agarose gels are usually run at a voltage of v V/cm.[iii] [6] The distance unit, cm, refers to the distance between the electrodes (i.e., the anode and the cathode) and not the length of the gel itself.[3] [6]
- Voltages too far below or above this range volition affect the mobility and the resolution of the bands. Low voltages will subtract the mobility and volition cause the bands to broaden. On the other hand, high voltages will subtract the resolution of the bands. This is largely due to the fact that voltages that are too high can cause the gel to overheat, and even melt.[eight]
- Concentration
- Agarose concentration must be taken into account when selecting a marker. The gel percent effects the migration of the DNA.[3] [half-dozen] Generally, the higher the gel concentration, the slower the rate at which the Deoxyribonucleic acid volition move through the gel. This is in addition to the office molecular weight plays in the migration of a DNA mark or sample, that is to say, that the higher the molecular weight, the slower the DNA will migrate.[3] [6]
- Gel concentration also affects the power to visualize the bands run out on the gel. Smaller bands are better resolved on a college percentage gel, whereas increased molecular-weight bands are more easily visualized on a lower per centum gel.[6]
Poly peptide markers [edit]
Development [edit]
Previously, protein markers had been adult using a variety of whole proteins. The evolution of a kit including a molecular-weight size mark based on poly peptide fragments began in 1993. This protein mark, composed of 49 dissimilar amino acrid sequences, included multidomain proteins, and allowed for the analysis of proteins cleaved at different sites.[9]
Current technique improvements in protein markers involve the employ of machine-development. The starting time auto-developed regularly-weight protein marker was invented in 2012.[10]
Design [edit]
Similar to Dna markers, these markers are typically composed of purified proteins whose molecular masses are already known.[3] The list beneath outlines some of the proteins, as well equally the molecular mass, that are commonly used when constructing a protein marker.

Immunoblot with protein ladder in leftmost lane. Fragment sizes are measured in kDA (kilodaltons).
| Protein | Molecular Mass (kDa) |
| Beta-galactosidase | 120[xi] |
| Phosphorylase B | 94[3] [12] |
| Bovine Serum Albumin (BSA) | 67[iii] [12] |
| Ovalbumin | 43[3] |
| Turkey Albumin | 40[12] |
| Carbonic Anhydrase | 30[iii] [12] |
| Soybean Trypsin Inhibitor | twenty.ane[3] [12] |
| a-Lactalbumin | 14.4[iii] [12] |
| Lysozyme | xiv[13] |
Choosing the correct poly peptide marker [edit]
Molecular-weight size markers can be broken up into ii categories: molecular weight markers vs. molecular ladder markers.[fourteen] Markers are either stained or unstained, and depending on the circumstance, one may exist more appropriate than some other. Molecular-weight size markers tin can likewise be biochemically contradistinct.[15] The conjugation with biotin is the near common. Molecular-weight size markers are most commonly used in SDS-polyacrylamide gel electrophoresis and western blotting. With all the different types and uses of molecular-weight size markers, information technology is of import to cull the advisable protein standard. Likewise the most mutual apply, as a style to summate the molecular weight of the samples, other uses include allowing visual evidence of protein migration and transfer efficiency and are sometimes even used for positive control.[16]
- MW marker vs Protein Ladders
- A molecular weight marker is one blazon of protein standard. They can either prestained or unstained prior to loading; depending on the blazon of experiment one may be more than advantageous. In either instance, they are normally run on the outer lane of a gel, while the sample is loaded in the middle lanes.[14] Molecular markers are different from protein ladders in that they are composed of a mixture of native proteins whose specifications are well categorized but practise not represent to whole numbers.[fourteen] Generally these are much cheaper, but analysis only allows for an guess value of the electrophoresis separated proteins.[14]
- A poly peptide ladder is another blazon of protein standard. They are almost always stained.[fourteen] Protein ladders differ from molecular markers in that they are composed of a mixture of highly purified proteins whose specifications are known and correspond to whole numbers.[fourteen] Mostly protein ladders are composed of x-12 proteins.[14] At the end of the experiment, later on size migration occurs, a single ring volition represent the size of each protein contained in the ladder.[17] Markers are evenly spaced, and size analysis using these markers allows for a precise value of the protein of involvement. In some cases, as a molecular confirmation method, MW markers are run with protein ladders for verification.[14]
- Prestained and Unstained Markers
- Protein markers tin come unstained or prestained, only both have their benefits and disadvantages.[18] Simple visualization of poly peptide separation and transfer is made possible through the use of prestained markers.[18] They are usually used in both SDS-polyacrylamide gel electrophoresis and western blotting. In SDS-PAGE information technology allows for the monitoring of protein migration, as the protein bands will split up and can be seen during an electrophoretic run. In western blots, the stained protein standards allow for the visualization protein transfer onto the membrane.[17] However, size determinations are non as accurate with these markers (encounter Recombinant and Natural Marker section for farther explanation).[18]
- While the unstained markers let for more exact size determinations, they cannot be viewed while the gel is running. Every bit such, the gel must be stained in order to visualize the bands.[xix]
- Recombinant and Natural Markers
- Too stained and unstained markers, poly peptide markers can be thought of in terms of recombinant and natural.[18] Recombinant markers consist of recombinant proteins which have been greatly purified. These markers are designed in such a way as to highlight particular characteristics.[18] Examples of these characteristics include affinity tags and molecular weights which are uniformly positioned relative to each other.[18]
- Natural markers, as the proper noun implies, are a mixture of proteins which happen naturally.[18] Prestained natural markers piece of work well for gel separation visualization. Yet, these markers tend to bind to the stain in a covalent manner in varying amounts and at various positions.[18] Consequently, the resultant bands may be broader. This is especially true when making comparisons to prestained recombinant markers. Due to this effect, molecular weight determinations are likely to exist less authentic with the prestained natural markers.[18]
- Biochemically Altered
- Protein standards can too be chemically altered. A mutual alteration is through the use biotin. Biotin has a very high affinity for streptavidin, and therefore, the bounden forms a very strong complex. For visualization, a colour tag is attached to the streptavidin.[15]
Effects of gel conditions [edit]
Every bit with Deoxyribonucleic acid electrophoresis, atmospheric condition such as buffers, charge/voltage, and concentration should be taken into account when selecting a protein marker.
- Buffers
- Buffers can affect the mobility of both the marker and the samples. The pH of the buffer varies with the organisation used and consequently, each buffer organization will have a dissimilar effect of the charge of a protein or proteins.[20] In addition, in the case of SDS-PAGE, the binding affinity for SDS can be affected by the buffering organization.[xx] Even when using the same percentage and type of gel, the same proteins will migrate at dissimilar rates depending on the buffer used.[xx]
- Charge/Voltage
- Voltage plays a part in the mobility of proteins on a gel. Proteins will migrate faster at college voltages. Consequently, the gel running time will be shorter. Conversely, higher voltages can result in greater band improvidence.[20] Also, if the voltage is as well high, the temperature in the electrophoresis sleeping accommodation can go such that the gel begins to melt.[20]
- The voltage that a gel should be run at depends on the blazon of the gel. For some gels, the voltage remains constant throughout the run, whereas, with other gels, the initial voltage is allowed to remain constant for a specified fourth dimension earlier it is increased.[20] This second voltage is so used for a specific time frame, after which, it may also be increased.[xx]
- Concentration
- In terms of percent, gels used for protein electrophoresis tin can be broken down into single-percentage gels and slope gels.[eighteen] Unmarried-per centum gels are likewise referred to as linear gels.[20] For linear gels, the selected percentage usually falls between 7.five% and 20%.[18] Common percentage ranges for gradient gels are four-15% and ten-xx%. Each type of gel has its own advantages.[xviii] For instance, linear gels are preferred when several proteins have similar molecular weights; better separation betwixt these proteins will be displayed by a linear gel.[18] On the other hand, gradient gels are a better option when the samples of involvement contain proteins of vastly different molecular weights or that cover a large range of molecular weights.[xviii] [20]
RNA markers [edit]
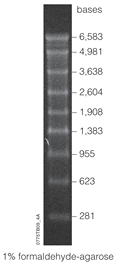
Lane of electrophoresed agarose gel showing an RNA ladder with bands at a range of 281-6583 base pairs
Development [edit]
RNA ladders composed of RNA molecular-weight size markers were initially developed by using the constructed circumvolve method[21] to produce different-sized markers. This technique was improved upon by inventor Eric T. Kool to use circular DNA vectors as a method for producing RNA molecular-weight size markers. As referred to as the rolling circle method, the improvements of this technique stems from its efficiency in synthesizing RNA oligonucleotides. From the round Dna template, unmarried-stranded RNA varying in length from 4-1500 bp can be produced without the demand for primers and by recycling nucleotide triphosphate. Dna can also be synthesized from the circular template, adding to this technique's versatility. In comparing to runoff transcription, the synthetic circle method produces RNA oligonucleotides without the runoff. In comparing to PCR, the synthetic circumvolve method produces RNA oligonucleotides without the need for polymerase nor a thermal cycler. This method is as well cost-efficient in its ability to synthesize grand amounts of product at a lower error rate than machine synthesizers.[21]
Design [edit]
The RNA markers consist of RNA transcripts of various incrementing lengths. For case, the Lonza 0.v-9 kbp marker[22] has bands marking 0.5, 1, 1.five, two, 2.v, three, four, five, 6, and 9 kilobase pairs. Markers are dissolved in a storage buffer, such as EDTA, and tin can have a shelf life of upward to 2 years when stored at -80 °C. To use the marker, such equally for northern blot analysis, it is first thawed, and so stained and so that information technology is detectable on a gel electrophoresis. One of the nigh mutual dyes used for markers is ethidium bromide.
The range of a detail marker refers to variety of bands it can map. A "high" range refers to relatively big fragments (measured in kb) while a "depression" range refers to markers that distinguish between pocket-sized fragments (measured in bp). Some markers can even be described equally "ultra-low range",[16] but even more precise is the microRNA marker. A microRNA marker can exist used to measure RNA fragments within a dozen nucleotides, such as the 17-25 nt microRNA marker.[23]
Use [edit]
At equivalent molecular weights, RNA will migrate faster than DNA. Notwithstanding, both RNA and Dna have a negative linear slope between their migration distance and logarithmic molecular weight.[24] That is, samples of less weight are able to migrate a greater distance. This relationship is a consideration when choosing RNA or Dna markers as a standard.
When running RNA markers and RNA samples on a gel, information technology is important to prevent nuclease contamination, as RNA is very sensitive to ribonuclease (RNase) degradation through catalysis.[25] [26] Thus, all materials to be used in the process must be taken into consideration. Any glassware that is to come into contact with RNA should be pretreated with diethylpyrocarbonate (DEPC) and plastic materials should be dispensable.[25]
Molecular-weight size markers and SDS-Page [edit]

SDS-PAGE gel using a molecular-weight size standard in the left outer lane
Ane of the nigh common uses for molecular-weight size markers is in gel electrophoresis. The purpose of gel electrophoresis is to divide proteins by physical or chemical properties, which include accuse, molecular size, and pH.< When separating based on size, the ideal method is SDS-PAGE or polyacrylamide gel electrophoresis and molecular-weight size markers are the advisable standards to utilise.
Gels tin can vary in size. The number of samples to be run will determine the appropriate gel size. All gels are divided into lanes that run parallel through the gel. Each lane will contain a specific sample. Typically, molecular-weight size standards are placed in an outer lane. If a gel has a specially high number of lanes, then multiple ladders may be placed across the gel for higher clarity.
Proteins and standards are pipetted on the gel in advisable lanes. Sodium dodecyl sulfate (SDS) interacts with proteins, denaturing them, and giving them a negative charge. Since all proteins have the same charge-to-mass ratio, protein mobility through the gel volition solely exist based on molecular weight. Once the electrical field is turned on, poly peptide migration will initiate. Upon completion, a detection machinery such as western blotting can exist used, which volition reveal the presence of bands. Each band represents a specific protein. The distance of travel is solely based on molecular weight; therefore, the molecular weight of each protein can exist determined by comparing the distance of an unknown protein to the standard of known molecular weight.[27]
Different uses of molecular-weight size markers [edit]
Many kinds of molecular-weight size markers exist, and each possess unique characteristics, lending to their involvement in a number of biological techniques. Option of a molecular-weight size marker depends upon the marker type (DNA, RNA, or poly peptide) and the length range it offers (e.one thousand. 1kb). Before selecting a molecular-weight size marking, it is important to become familiar with these characteristics and properties. In a particular instance one type may be more advisable than another. Although specific markers can vary between protocols for a given technique, this section will outline general markers and their roles.
Allozymes [edit]
The first blazon of molecular marker developed and run on gel electrophoresis were allozymes. These markers are used for the detection of protein variation. The word "allozyme" (also known as "alloenzyme") comes from "allelic variants of enzymes."[28] When run on a gel, proteins are separated by size and charge. Although allozymes may seem dated when compared to the other markers available, they are all the same used today, mainly due to their low cost. 1 major downside is that since there is only a limited amount bachelor, specificity an issue.[28]
Dna-based markers (1960s) [edit]
Although allozymes can notice variations in DNA, information technology is past an indirect method and not very accurate. DNA-based markers were developed in the 1960s.[28] These markers are much more effective at distinguishing between Dna variants. Today these are the most commonly used markers. DNA-based markers work by surveying nucleotides, which can serve a diversity of functions, such as detecting differences in nucleotides or even quantifying the number of mutations.[28]
- RFLP
- Restriction fragment length polymorphism is a technique used to detect variations in homologous DNA.[29] Specific restriction endonucleases are used to digest Dna. The RFLP molecular mark is specific to a unmarried fragment. Forth with alleic RFLP markers, a molecular-weight size marking, in this case a DNA marker,[30] is likewise included on an electorphoresed agarose gel. The Deoxyribonucleic acid marker allows for the size of the restriction fragments to be estimated.
- Minisatellites
- Like to RFLP, this technique too uses brake endonucleases to digest the genomic DNA. Minisatellites are short sequences of tandem repeats, approximately x-threescore base of operations pairs. Minisatellites can be used in Dna footprinting and every bit regulators of cistron control.[28]
PCR-based markers (1980s) [edit]
The success of Deoxyribonucleic acid based markers lead to the development of PCR. PCR (polymerase chain reaction) is a DNA amplification technique that can be practical to various types of fragments. Prior to this development, to amplify Dna, it had to be cloned or isolated. Shortly after the discovery of PCR came the idea of using PCR-based markers for gel electrophoresis. These type of markers are based on PCR primers and are categorized as Deoxyribonucleic acid sequence polymorphism.[28]
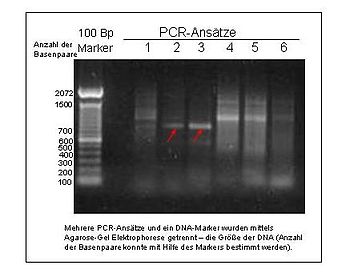
An electrophoresed gel showing PCR products. The leftmost lane represents a DNA ladder with fragments at 100bp intervals.
- Microsatellites
- Also known every bit SSR (uncomplicated sequence repeats) or STR (short tandem repeats), microsatellites differ from minisatellites in that they are shorter, usually 2-6 base pairs. This belongings of microsatellites allows for easy isolation. Microsatellites are most commonly used in population genetics. Microsatellites have a loftier and complex mutation rate, which is their major disadvantage.[28]
- AFLP
- Amplified fragment length polymorphism is a PCR-based DNA fingerprinting technique. DNA is showtime digested with endonucleases. The brake fragments are then ligated together.[31] A molecular marker is then generated when specific fragments are selected for amplification. AFLP markers are run alongside a DNA marking on a gel. A common AFLP DNA mark is xxx-330bp long.[32] The fragments of this mark prevarication at 10bp intervals to increase precision.
- RAPD
- Random amplified polymorphic DNA is a technique that is conducted similar to AFLP. The difference is that the molecular markers are generated at random.[31] The most common molecular-weight size mark for this technique is the 1kb DNA ladder.[33] [34]
DNA sequence polymorphism [edit]
Although technically speaking, DNA sequence polymorphism has been going on since the use of RFLP in the 1960s, the analysis has changed significantly over the years. Dna sequence polymorphism uses older techniques similar RFLP, just on a larger scale. Sequencing is much faster and more efficient. The analysis is automated, as it uses a technique known equally shotgun sequencing. This loftier-throughput method is commonly used in population genetics.[28]
- SNPs
- SNPs (unmarried nucleotide polymorphism), are used to observe variations in single nucleotides. The technique is very like to that of RFLP. SNPs are used frequently for population genetic studies.[35] Later amplification through PCR, these small fragments tin can exist visualized using gel electrophoresis, and over again Dna markers play a role in determining fragment length.
Polysaccharide analysis by carbohydrate gel electrophoresis [edit]
Carbohydrate markers are employed in a technique known every bit polysaccharide analysis by saccharide gel electrophoresis (PACE), which is a measurable separation technique.[36] It allows for the analysis of enzyme hydrolysis products.[36] It has been used in applications such as characterizing enzymes involved in hemicellulose degradation, determining the structure of hemicellulose polysaccharides, and assay of enzymatic cleavage of cellulose products.[36]
PACE depends on derivitization, which is the conversion of a chemic compound into a derivative.[36] [37] Hither monosaccharides, oligosaccharides, and polysaccharides are the compounds of interest. They are labeled at their reducing ends with a fluorescent characterization (i.east. a fluorophore).[36] This derivitization with a fluorophore permits both separation on a gel under the desired circumstances and fluorescence imaging of the gel. In this case, a polyacrylamide gel is used.[36]
Every bit with DNA, RNA, and protein electrophoresis, markers are run alongside the samples of interest in carbohydrate gel electrophoresis.[36] The markers consist of oligosaccharides of known molecular weight. Like the samples of interest, the mark is besides derivitized with a fluorophore (usually with 8-aminonaphthalene-1,3,6-trisulfonic acid (ANTS) or 2-aminoacridone).[36]
References [edit]
- ^ Carlson, David P. "Size markers for electrophoretic analysis of Dna". Usa Patent #5316908A. Google Patents. Retrieved 30 October 2013.
- ^ Carlson, David P. "Size markers for electrophoretic analysis of DNA". EP Patent #0466404B1. Google Patents. Retrieved 30 October 2013.
- ^ a b c d eastward f g h i j thousand fifty 1000 n o p Blaber, Mike. "Lecture 20: Gel Electrophoresis". BCH5425 Molecular Biology and Biotechnology.
- ^ Bowen, R (20 Oct 1999). "DNA Ligation". Biotechnology and Genetic Applied science . Retrieved 12 Nov 2013.
- ^ a b c Lan, Vo Thi Thuong; Loan, Pham Thi Thanh; Duong, Pham Anh Thuy; Thanh, Le Thi; Ha, Ngo Thi; Thuan, Ta Bich (2012). "Straightforward Procedure for Laboratory Production of Deoxyribonucleic acid Ladder". Journal of Nucleic Acids. 2012: 254630. doi:ten.1155/2012/254630. PMC3306978. PMID 22496965.
- ^ a b c d eastward f yard Bowen, R. (2000). "Agarose Gel Electrophoresis of DNA". Hypertexts for Biomedical Sciences – Colorado State Academy.
- ^ "Tris Borate EDTA and Tris-Acetate-EDTA Buffer (TAE & TBE, pH viii.3)" (PDF). Aniara.
- ^ a b c "Agarose Gel Electrophoresis Tips & Tricks". Life Technologies.
- ^ Hartley, James. "Protein size marker ladder". US Patent #5449758A. Google Patents. Retrieved thirty Oct 2013.
- ^ Cheng, Tian Lu. "Auto-developing and regularly-weighted poly peptide molecular weight marker kit and method for preparing the same". U.s.a. Patent #20130217133A1. Google Patents. Retrieved xxx October 2013.
- ^ "Prestained Poly peptide Molecular Weight Marker". ThermoScientific . Retrieved 12 Nov 2013.
- ^ a b c d e f Ingelman, Margareta (2004). "Poly peptide separation and analysis". KE7001 Biochemistry Labs . Retrieved 12 November 2013.
- ^ "Protein Molecular Weight Markers". Aid Biotech. 2011. Retrieved 12 November 2013.
- ^ a b c d east f 1000 h "Protein Molecular Weight Markers Comparing and Choice Guide". Retrieved 16 November 2013.
- ^ a b "Biotinylated Molecular Weight Marker". Retrieved xvi November 2013.
- ^ a b "Molecular Weight Markers". Retrieved 16 November 2013.
- ^ a b "Pierce Prestained Protein Molecular Weight Marker". Retrieved 16 November 2013.
- ^ a b c d e f 1000 h i j k l yard n "Electrophoresis A Guide to Polyacrylamide Gel Electrophoresis and Detection" (PDF). Bio-Rad.
- ^ "Poly peptide Molecular Weight Markers Comparison and Pick Guide". ThermoScientific . Retrieved 12 November 2013.
- ^ a b c d due east f g h i "Protein Handbook 2013" (PDF). Life Technologies.
- ^ a b Kool, Eric T. "Round Deoxyribonucleic acid vectors for synthesis of RNA and Dna". US Patent #6096880A. Google Patents. Retrieved 27 November 2013.
- ^ Lonza. "RNA Markers 0.5–9 kbp" (PDF). Document #18123-0807-06. Lonza Rockland Inc. Retrieved 27 November 2013.
- ^ New England Biolabs. "microRNA Mark". New England Bioloabs. Retrieved 27 November 2013.
- ^ Wicks, Richard J. (1986). "RNA molecular weight conclusion by agarose gel electrophoresis using formaldehyde as denaturant: Comparing of rna and deoxyribonucleic acid molecular weight markers". International Journal of Biochemistry. xviii (3): 277–278. doi:x.1016/0020-711x(86)90118-7. PMID 2937672.
- ^ a b "Itemize # : R0004". RNA Marker High Like shooting fish in a barrel. Abnova. Retrieved xiv December 2013.
- ^ "RNA Electrophoresis: Introduction". RNA Electrophoresis. Thermo Fisher Scientific Inc. Retrieved 14 December 2013.
- ^ "Molecular Weight Determination of Proteins" (PDF) . Retrieved fourteen December 2013.
- ^ a b c d east f g h Schlotterer, Christian. "The Evolution of Molecular Markers" (PDF) . Retrieved 26 November 2013.
- ^ "Population Improvement". Retrieved thirty October 2013.
- ^ Higgins, Fifty. (Apr 2012). "Deoxyribonucleic acid Ladder for Gel Electrophoresis". Lewis & Clark College. Retrieved xv Nov 2013.
- ^ a b Mueller, Ulrich (1999). "AFLP genotyping and fingerprinting" (PDF). Trends in Ecology & Evolution. 14 (10): 389–394. doi:10.1016/s0169-5347(99)01659-6. PMID 10481200. Retrieved 30 October 2013.
- ^ Invitrogen Corporation (2003). "30-330 bp AFLP® Deoxyribonucleic acid Ladder" (PDF). Manual. Life Technologies Corporation. Retrieved 15 Nov 2013.
- ^ Gianniny, Christine; et al. (May 2004). "unRAPD analysis of mtDNA from tomato flowers free of nuclear DNA artifacts" (PDF). BioTechniques. 36 (5): 772–776. doi:10.2144/04365BM04. PMID 15152595. Retrieved 15 Nov 2013.
- ^ Roberts, M. A.; Crawford, D. L. (1 June 2000). "Use of Randomly Amplified Polymorphic DNA as a Means of Developing Genus- and Strain-Specific Streptomyces Deoxyribonucleic acid Probes". Applied and Environmental Microbiology. 66 (6): 2555–2564. doi:10.1128/AEM.66.half-dozen.2555-2564.2000. PMC110581. PMID 10831438.
- ^ McClean, Phillip. "The Classes of Molecular Markers". Retrieved xxx October 2013.
- ^ a b c d eastward f thousand h Kosik, Ondrej; Bromley, Jennifer R.; Busse-Wicher, Marta; Zhang, Zhinong; Dupree, Paul (2012). "Studies of Enzymatic Cleavage of Cellulose Using Polysaccharide Analysis past Saccharide gel Electrophoresis (Stride)". Methods in Enzymology. 510: 51–67. doi:10.1016/B978-0-12-415931-0.00004-five. ISBN9780124159310. ISSN 0076-6879. PMID 22608721.
- ^ Cammack R, Attwood TK, Campbell PN, Parish HJ, Smith A, Stirling JL, Vella F (2006). "Fluorophore". Oxford Dictionary of Biochemistry and Molecular Biology (2d ed.). Oxford University Press.
Source: https://en.wikipedia.org/wiki/Molecular-weight_size_marker
0 Response to "How to Read Dna Molecular Weght Markers"
Post a Comment